

Table of Contents
- Summary & Introduction
- Epifaunal Worm Tubes on Lower Lias Ammonites – Results
- Epifaunal Worm Tubes on Lower Lias Ammonites – Detailed Observations and interpretation
- Epifaunal Worm Tubes on Lower Lias Ammonites – Discussion
- Epifaunal Worm Tubes on Lower Lias Ammonites – Conclusions and References
5. Discussion
5.1. Path of the worm tubes
From the worm’s point of view, successful attachment required that the ammonite survived for long enough for the worm to reach maturity and breed. This was unlikely if, for example, attachment occurred shortly before the death of the ammonite. We have several ammonites with very short worm tubes attached, some of which may have died prematurely due to the death of the ammonite shortly after their attachment to it. Attachment to juvenile ammonites was preferable.
Modern serpulid worms deploy two branchia on either side of the aperture of the worm tube. The backward-sloping orientation of the worm tube aperture facilitates raising the branchia above the substrate. Fossil examples where the initial attachment was in the umbilicus of the ammonite would raise both branchia on the same side of the ammonite. Growth of worm tubes around the venter is a more effective position for the worm as it could deploy its branchia on either side of the ammonite. It may also have been less of a hindrance to the ammonites as the drag would have been symmetrical.

5.1.1. Site of initial attachment
Grooves seem to have been the larval worm’s preferred sites of attachment. In some cases the worm tube continued all its growth along the umbilical seam (Fig. 12). Furthermore, worm tubes growing radially towards the periphery typically grew in the troughs between the ammonite’s ribs (e.g., Fig. 7). These observations suggest complex behaviour on the part of the worm in selecting an initial site of attachment and then growing in a manner that minimized the inconvenience to the ammonite host.
Fig. 12. Left side of LYMPH 2010/46 a Promicroceras with an epifaunal worm tube that grew entirely along the umbilical seam. Scale bar = 5 mm.
5.1.2. Subsequent growth
After initial attachment the worm attempted to grow so that its aperture remained in the ideal position with respect to the currents generated by the ammonite. Worms that grew entirely within the umbilical seam (Fig. 12) attached to juvenile ammonites and subsequently kept pace with the growth of the ammonite, thus maintaining the ideal position for their apertures. We presume that the growth of a worm tube radially across a whorl was caused by the worm tube effectively growing faster (as measured in degrees) than the ammonite. Thus, the worm tube aperture would start to approach the less ideal positions at 3 or 9 o’clock (Fig. 8).
5.1.3. Position of the worm tube aperture relative to the ammonite
Regrettably, in none of our examples can we demonstrate that the ammonite shell is complete. However, the worm tubes did not reach the ammonite aperture in life (Figs 2-7). One example of Schindewolf’s (1934, pl. 2, figs 4a, b) shows the worm tube aperture approximately 115o behind the aperture of Schlotheimia angulosa Lange. Hence, the worm tube aperture was near the lowest point on the shell with the ammonite in its normal life position (Trueman 1941, fig. 14). His illustration shows the ammonite aperture horizontal and a body chamber of approximately 240o. We estimate that for our examples of Promicroceras with worm tubes attached the worm tube aperture lay about 105o behind the ammonite aperture, which is close to the value on Schindewolf’s example of Schlotheimia (Fig. 8D).
From all these sources it seems that the worm tube aperture was close to the lowest point on the ammonite shell in life orientation. The worm tubes apparently ended about 105-115o behind the ammonite aperture. Hence, most of their additional mass would have weighed down part of the ammonite rotating the shell clockwise (as illustrated in Figs 8 and 9) and raising the aperture even further. Furthermore, deconstructing Schindewolf’s original example on the assumption that the worm tube grew at the same rate as the ammonite implies that the initial point of attachment of the worm tube was just over 90o back from the aperture (Fig. 8A). If Schindewolf’s example also held its aperture horizontal, this means the initial attachment was near the lowest point on the umbilical seam. We assume that initially the worm grew along the umbilical seam and kept pace with the ammonite (Fig. 8B). It then grew across what was at the time the outer whorl because it began to grow forward faster than the ammonite (as measured in degrees) and so its aperture started to climb up from 6 o’clock (the ideal position) towards 9 o’clock (Fig. 8C). Modern serpulids grow faster as juveniles than as adults (Kupriyanova et al., 2001). On reaching the venter of the ammonite, the worm apparently kept pace with ammonite growth (Fig. 8D).
In Promicroceras along any radial line the distance from the origin to the umbilical seam is almost exactly half the distance from the origin to the venter (Fig. 8D). Thus, for any fixed angle of growth the linear increase in length along the umbilical seam is almost exactly half that along the venter (Fig. 8D). Worm tubes lying in the umbilical seam that kept pace with ammonite growth grew at half the pace (as measured in mm) of those that grew on the venter. Although we have three examples in which all the growth of the worm tube is primarily along the umbilical seam, most sooner or later crossed a whorl to gain the venter. We assume that this was because they began to grow at a faster angular rate than the ammonite while in the umbilical seam and that moving across the outer whorl allowed the ammonite to catch up. All this points to the fact that most worm tubes that attached to juvenile ammonites did so close to 6 o’clock as the ammonite was orientated at the time and subsequently they did all they could to maintain their apertures in this position.
5.2. Ecology of attachment
In most of our examples the worm tubes first attached to the ammonites when the latter were juvenile. Chapman et al. (2007) reported a significant decline in settlement of the Recent worm Serpula vermicularis in Loch Creran, Scotland, with increasing depth between 6 and 12 m. Cotter et al. (2003) recorded greater recruitment at 7 m than at 4 m in Bantry Bay, Ireland, for two species of Pomatoceras and one of Serpula. Interestingly, the latter also demonstrated greater larval settling on the undersides of experimental substrates compared to the upper surfaces. Recent serpulids seem to have preferred depths and sites for larval settlement. We wonder if juvenile ammonites lived in shallower water than adults and thus may have been more exposed to serpulid larval settlement.
If the worm tubes kept pace with the growth of the ammonite shell, one can speculate about the rate of growth of the ammonites. Schindewolf (1934) concluded that growth of the last whorl of the ammonites he illustrated took between six months and three years, based on previously published growth rates of five species of Recent serpulids. Since then a great deal more has been published on worm tube growth rates (see the review in Kuprianova et al., 2001). In particular, Kuprianova et al. recorded substantial changes in growth rates with age (growth rates are fastest in the earliest growth stages). Thus, short-term growth rates are unreliable for estimating growth rate in fossils. However, Hill (1967) and Medernach et al. (2000) showed that modern serpulids reach 30-40 mm tube length in four months to one year, but live for one to two years with little or no subsequent growth. If one accepts a growth rate of about 30 mm in a year, this is equivalent to the length of the periphery of a Promicroceras about 10 mm in diameter, and is about as long as our worm tubes ever grew. Thus, we think it reasonable to assume that Promicroceras reached adult size in two to three years and may well have lived for considerably longer after becoming fully grown.
5.3. Relationship between the ammonites and worm tubes
All the Promicroceras with epifaunal worm tubes are small, although some are wholly septate and incomplete. Nevertheless, some died young and the attachment of worm tubes may have contributed to their deaths. We have used the diameter of the ammonites at the last suture as a measure of size that is not affected by fossils having incomplete body chambers. Mean diameter of Promicroceras with worm tubes is 11.04 mm (n = 25), whereas mean diameter of fatally bitten Promicroceras without worm tubes is 16.3 mm (n = 15) (Andrew et al., 2010). Thus, whether adult or juvenile, Promicroceras with worm tubes were smaller when they died than those without. We also measured the width and height of the ammonite whorl at the last suture, as well as the width and height of the worm tubes at their apertures to investigate the effect of the extra mass the worm tubes must have added to the ammonites. We have used the ratio of the sum of the cross-sectional areas of all worm tubes attached to one ammonite, divided by the cross-sectional area of that ammonite at its last suture, as a measure of the ammonite’s ability to maintain buoyancy hampered by this additional mass. A highly significant inverse correlation exists between the diameter of the ammonites at death and this ratio (Fig. 13). Ammonites with larger or with more worm tubes died at smaller sizes. From both sets of data, the presence of the worm tubes was harmful to the ammonites and the relationship was parasitic.
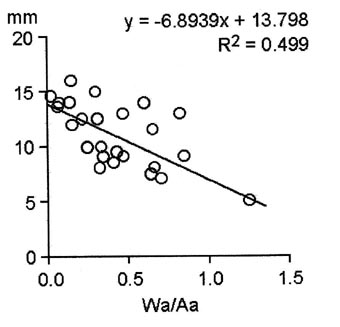
Fig. 13. Diameter of ammonites at the final suture (mm) versus estimated additional weight of worm tubes, measured as the ratio of total cross-sectional area of worm tubes to cross-sectional area of the ammonite at its final suture (Wa/Aa). The negative correlation implies that the higher the ratio, the smaller the size at which the ammonites died. The correlation coefficient is highly significant (p<.001).
Attachment of worm tubes was not always fatal. One example, a Microderoceras (LYMPH 2010/64) from the Birchi Nodular Bed (base of Bed 75) survived the attachment of worm tubes. It has at least three worm tubes growing along the venter (Fig. 14). This specimen is larger than most of the others. As preserved it reaches 63 mm diameter and was much larger originally. It coiled normally to about 7.5 mm diameter where there is a slight deformity to the whorl, which started to grow away from the preceding whorl due to the presence of the worm tubes. Three worm tubes can be seen to have grown for about half a whorl and they too grew up away from the underlying ammonite venter. All three worm tubes then ended abruptly, but the ammonite grew back down to regain contact with the preceding whorl and continued to grow normally, except that the irregularity in growth caused by the presence of the worm tubes was repeated in the succeeding two whorls. At the extreme of the distortion a 3 mm gap exists between the ammonite whorls. At this point the ammonite was only 16 mm across, so the distortion is severe. The lowest of the three worm tubes is also 1.25 mm clear of the underlying ammonite venter, a distance equal to about twice its internal diameter as preserved. It would appear that the worm tubes themselves were overgrowing something attached to the ammonite, which is not now preserved.

Fig. 14. LYMPH 2010/64. A, left side of a Microderoceras from the Birchi Nodular Bed to show the distortion caused by the presence of worm tubes. B, detail of the worm tubes (1-3). Scale bars = 5 mm.
Finally, we have assumed that the benefit to the worms was the increased food-gathering capacity produced by the currents generated by the swimming ammonite. In several of our worm tubes the angle of the growth lines increases just before the aperture and one (LYMPH 2010/59) bears two successive thickenings of the aperture. Both points suggest that these worm tubes were mature. This is even true of some secondary worm tubes adjacent to the primary worm tube on the venter (e.g., LYMPH 2010/53; 2 and 3 in Fig. 5A). So, although the secondary worms were smaller than the primary worms, apparently they were still mature. If these interpretations are correct, the worms were able to reach maturity and (presumably) breed before the ammonites succumbed to the effects of their extra weight. This interpretation also tends to confirm that Promicroceras lived for some time after reaching full size. If the modified apertures indicate that the worm tubes had stopped growing yet the worms normally attempted to keep their apertures at the ‘ideal’ 6 o’clock position, this implies that the Promicroceras had also ceased growing. Again, modification of apertures is not normally cost effective if the animal, whether worm or ammonite, dies immediately. Thus, modified worm tube apertures indicate that both the worms and the ammonites potentially lived for some time without increasing in size.